【PBJ】茉莉酸信号在番茄冷胁迫反应中的调控机制 ——MYB15-LOXD 和 MYB15-MYC2-LOXD 模块的解析

【遗传转化项目进展汇报】
2025 年 7 月 15 日
【Advanced Science】里程碑突破:GmSop20——破解大豆油蛋白负相关的”平衡基因”
2025 年 7 月 25 日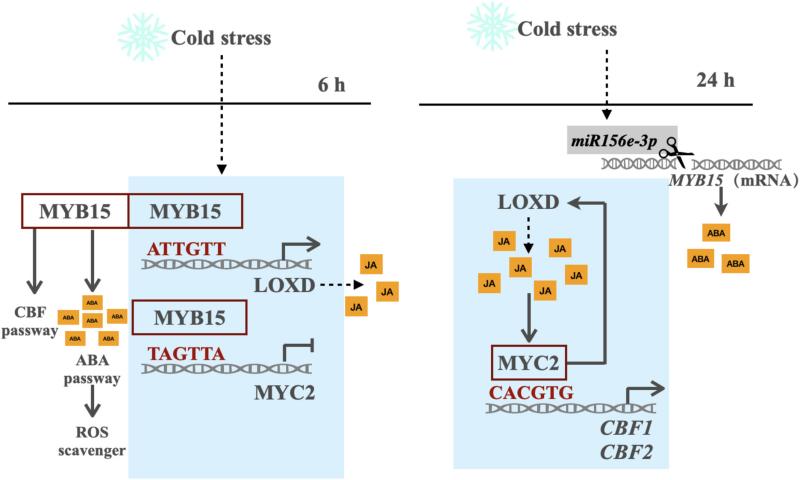
2025年6月份西北农林科技大学李建明教授在Plant Biotechnology Journal上发表了题为Regulation of jasmonic acid signalling in tomato cold stress response: Insights into the MYB15-LOXD and MYB15-MYC2-LOXD regulatory modules的研究论文,该研究揭示了番茄在冷胁迫下,SlMYB15 在初期激活 SlLOXD 促进 JA 合成并抑制 SlMYC2,后期 SlMYB15 表达下降,SlMYC2 激活 SlLOXD 和 SlCBF1/2 以维持耐寒性的动态调控机制。这一发现阐明了 MYB15-LOXD 和 MYB15-MYC2-LOXD 模块在番茄冷胁迫响应中的关键作用,为提高作物抗寒性提供了理论依据。
番茄是全球重要的经济作物,但冷胁迫会严重抑制其生长发育并降低产量。植物通过复杂的信号网络应对冷胁迫,其中激素信号(如 ABA、茉莉酸 JA)是关键调控因子。已知 ABA 在冷胁迫初期(6 小时)快速积累,但其持续高水平会抑制生长;而 JA 的作用机制,尤其是在冷胁迫中后期的调控模式,尚未明确。
本文聚焦于 JA 信号在番茄冷胁迫响应中的动态调控,重点探究转录因子 SlMYB15、JA 合成关键基因 SlLOXD 及 JA 信号核心因子 SlMYC2 之间的相互作用,旨在揭示 “MYB15-LOXD” 和 “MYB15-MYC2-LOXD” 两个调控模块如何协同调控番茄的耐寒性。
核心发现概述
研究发现,番茄在冷胁迫(4℃)下,JA 信号与 ABA 信号呈现 “时间接力” 模式:ABA 在 6 小时达峰,而 JA 在 24 小时达峰。这一过程由 SlMYB15 主导的动态调控实现:
冷胁迫初期(0-6 小时),SlMYB15 激活 SlLOXD 表达以促进 JA 合成,同时抑制 SlMYC2 表达,优先启动 ABA 通路;
冷胁迫中后期(6-24 小时),SlMYB15 表达受 miRNA 抑制而下降,对 SlMYC2 的抑制解除,SlMYC2 接手激活 SlLOXD 和 SlCBF1/2,维持 JA 信号并增强耐寒性。
关键结果解析
1.冷胁迫下 ABA 与 JA 的动态积累
ABA:在 4℃处理 6 小时达峰(较对照增加 4.28 倍),随后逐渐下降,体现 “快速响应、短期作用” 特征。
JA:在 24 小时达峰(较对照增加 3.47 倍),持续时间更长,暗示其在冷胁迫中后期起主导作用。
这一 “时间差” 为两种激素的功能分工提供了基础:ABA 应对急性冷胁迫,JA 维持长期耐寒性。
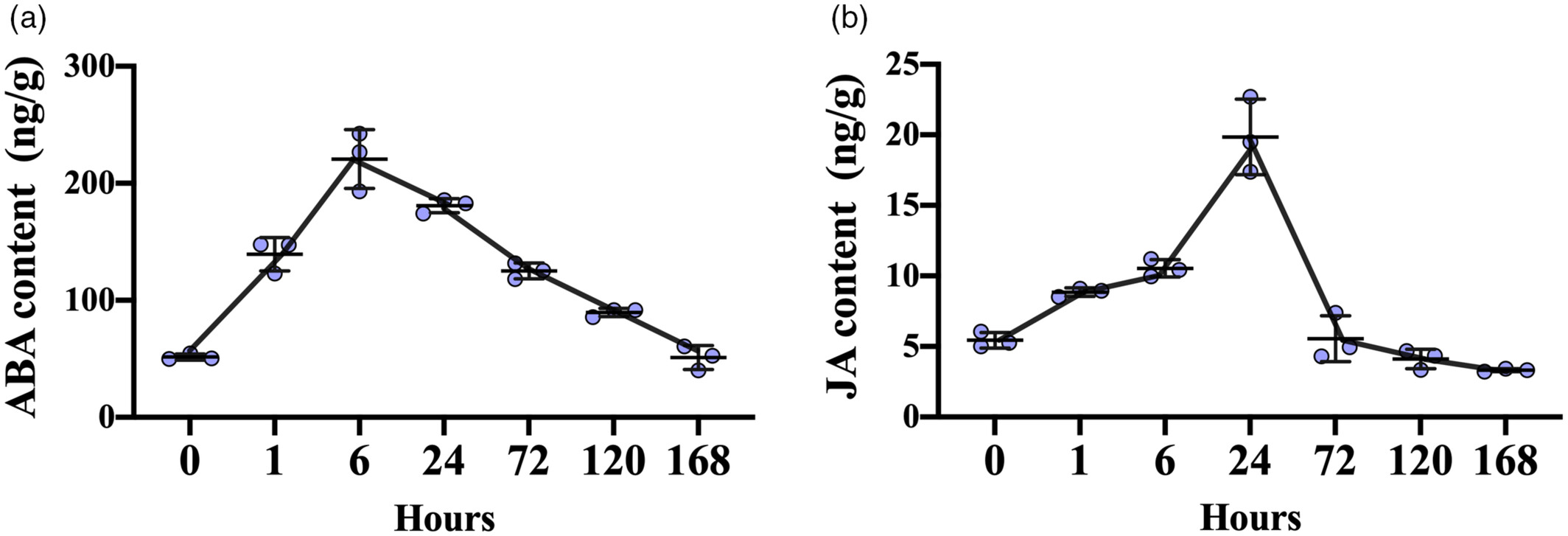
图1寒冷胁迫诱导番茄叶片脱落酸和茉莉酸积累
2.SlLOXD 是 JA 合成与耐寒性的核心调控因子
SlLOXD 的表达模式:在叶片、花、果实中高表达,冷处理 24 小时达峰,与 JA 积累趋势一致。
功能验证:
过表达 SlLOXD(LOXDOE lines):JA 含量升高,叶片相对电解质渗漏率降低,光系统 II 活性(Fv/Fm)提高,耐寒性增强。
突变体(loxd-6/7):JA 含量下降,耐寒性显著减弱。
结论:SlLOXD 通过促进 JA 合成直接调控番茄耐寒性。
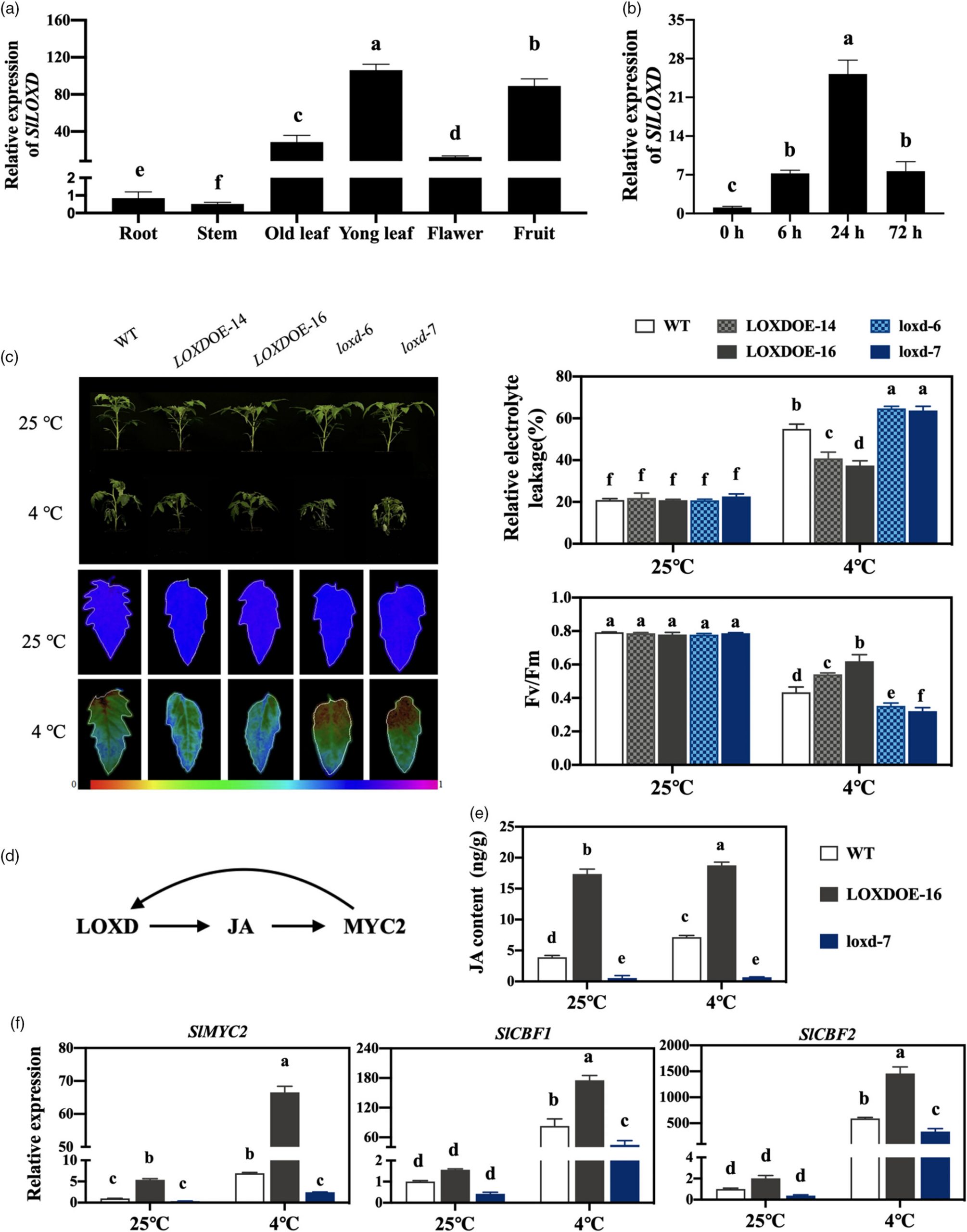
图2 SlLOXD 在调节番茄的耐寒性中发挥作用
3. SlMYC2 通过激活 SlCBF1/2 增强耐寒性
SlMYC2 的作用:作为 JA 信号的核心转录因子,直接结合 SlCBF1/2 的启动子(通过 EMSA 和双荧光素酶实验验证),激活其表达。
功能验证:
沉默或突变 SlMYC2(TRV2-SlMYC2、slmyc2-13):SlCBF1/2 表达下降,叶片枯萎严重,电解质渗漏率升高,耐寒性降低。
过表达 SlLOXD 的植株中,SlMYC2 表达同步上调,印证 SlMYC2 与 JA 信号的关联性。
结论:SlMYC2 通过调控 SlCBF1/2(冷响应核心基因)传递 JA 信号,增强耐寒性。
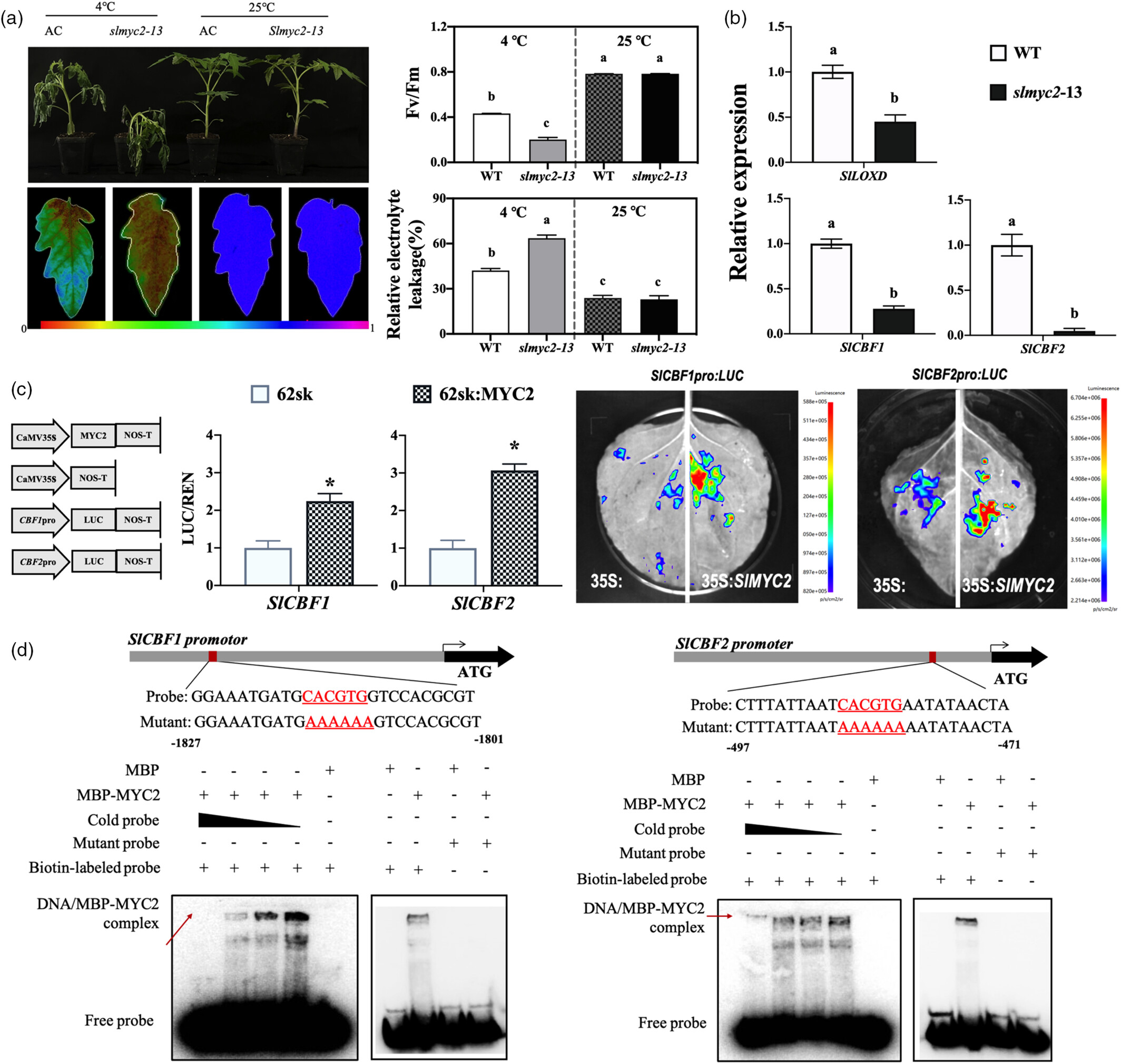
图3 SlMYC2 转录调节 SlCBF1/2 并参与番茄的耐寒性
4.SlMYB15 在冷胁迫初期激活 SlLOXD
SlMYB15 与 SlLOXD 的相互作用:
体内(ChIP-qPCR)和体外(EMSA、Y1H)实验证明,SlMYB15 直接结合 SlLOXD 启动子的 TAACAA 基序,激活其表达。
过表达 SlMYB15(MYB15OE-18):SlLOXD 表达升高 5.63 倍,JA 含量显著增加,耐寒性增强。
时间特异性:冷胁迫 6 小时内 SlMYB15 表达高峰,与 SlLOXD 的初期激活一致,说明其在 JA 合成启动阶段的关键作用。
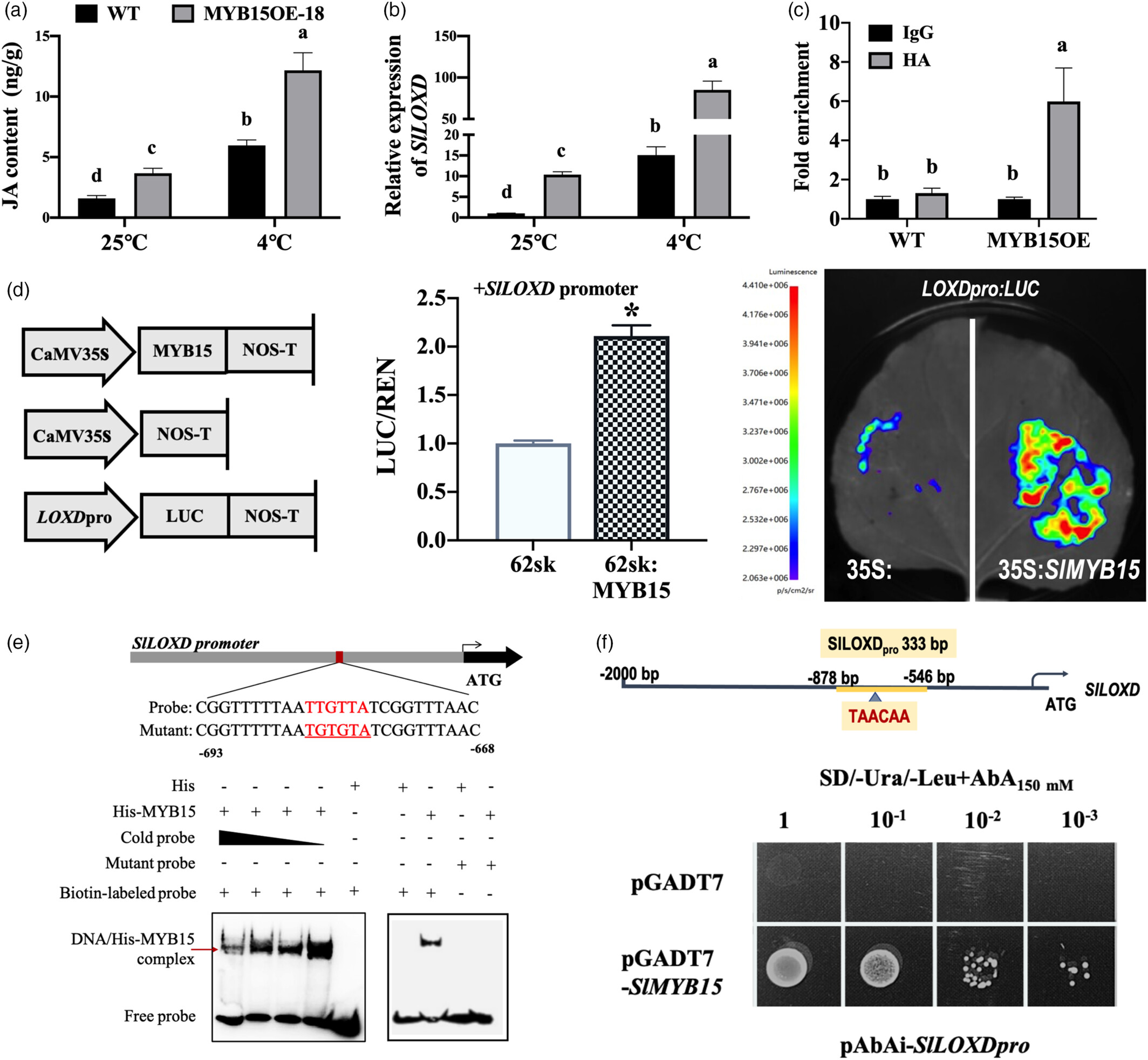
图4 SlMYB15 通过 SlLOXD 的转录调控增强冷处理下的 JA 积累
5.SlMYB15 对 SlMYC2 的抑制与调控接力
SlMYB15 抑制 SlMYC2:
SlMYB15 直接结合 SlMYC2 启动子的 TAGTTA 基序(EMSA、Y1H 验证),抑制其表达(双荧光素酶实验显示 LUC 活性降低)。
过表达 SlMYB15 的植株中,SlMYC2 表达显著下调,印证抑制关系。
调控接力机制:
6 小时后,SlMYB15 因 miR156e-3p 抑制而表达下降,对 SlMYC2 的抑制解除,SlMYC2 表达上调。
SlMYC2 对 SlLOXD 的激活作用更强(双荧光素酶实验显示其诱导的 LUC 活性高于 SlMYB15),维持 SlLOXD 高表达和 JA 合成。
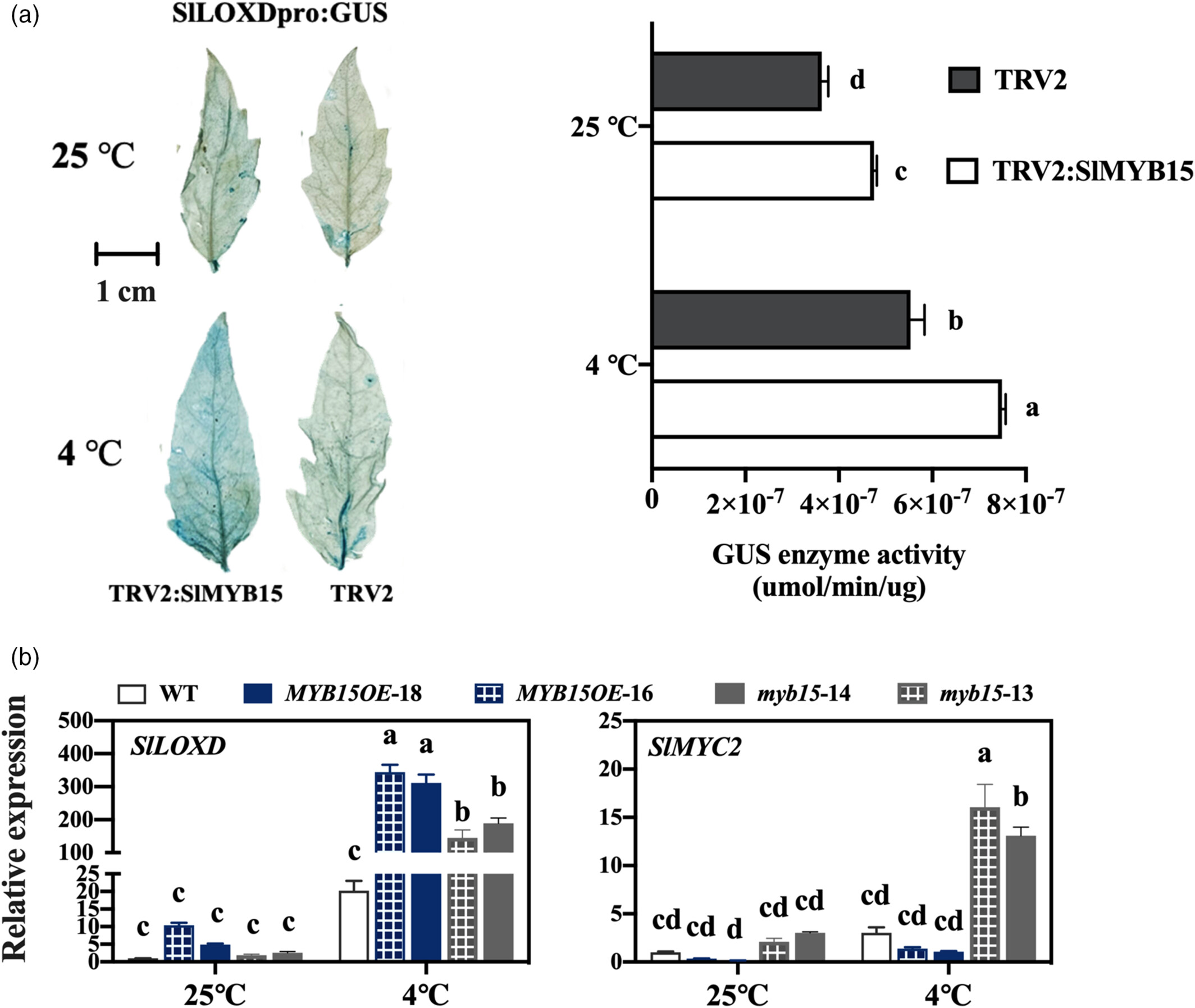
图5 SlLOXD 在 SlMYB15 沉默的番茄和 slmyb15 突变体 (myb15-14/13) 的叶片中上调
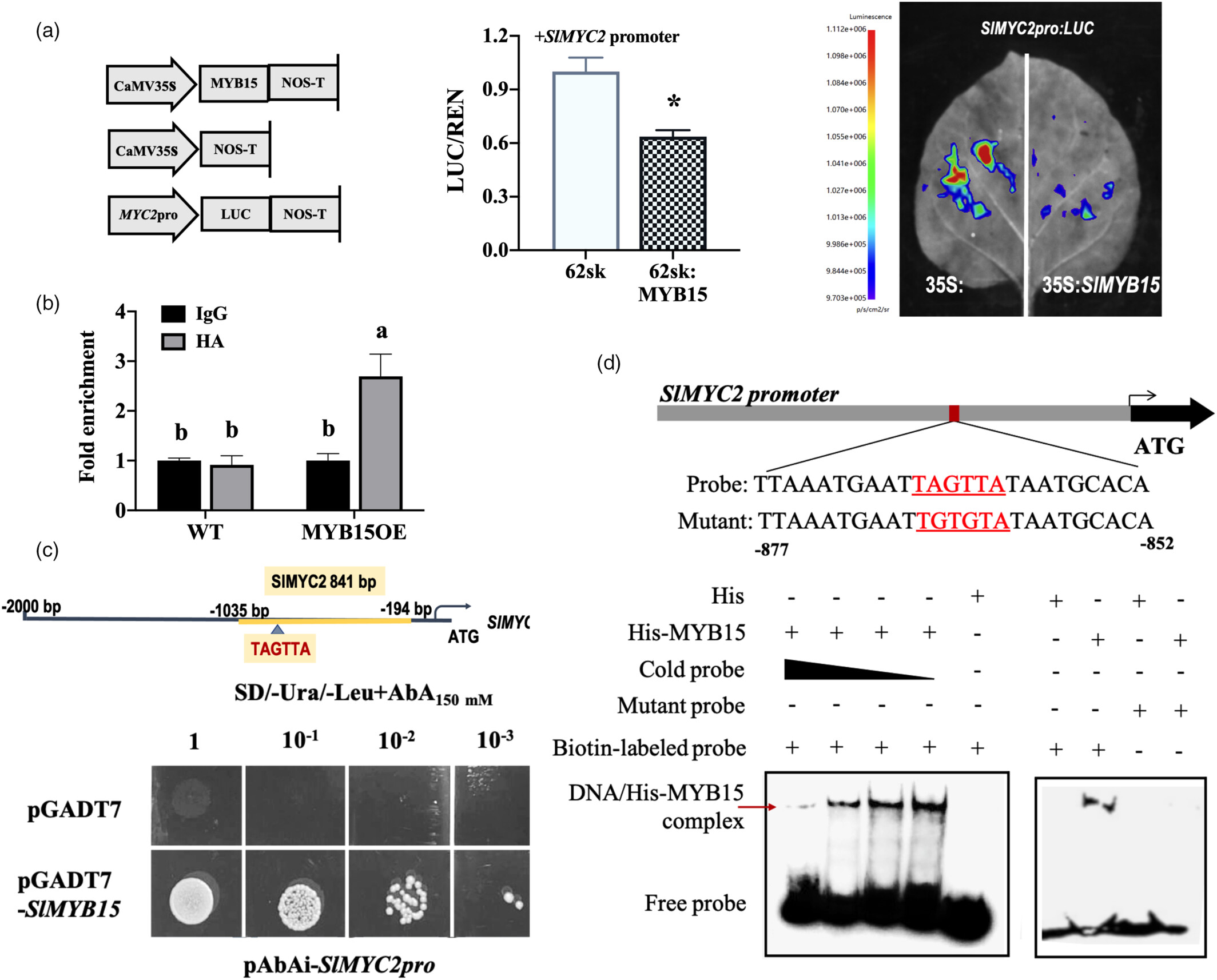
图6 SlMYB15 负向调节 SlMYC2 启动子活性
6.模块协同作用:MYB15-LOXD 与 MYB15-MYC2-LOXD 的动态平衡
冷胁迫 0-6 小时:SlMYB15 主导,激活 SlLOXD 以启动 JA 合成,同时抑制 SlMYC2,确保 ABA 通路优先响应。
冷胁迫 6-24 小时:SlMYB15 表达下降,SlMYC2 释放并激活 SlLOXD 和 SlCBF1/2,形成 “SlMYC2-SlLOXD” 正反馈,维持 JA 信号和耐寒性。
共沉默 SlMYB15 和 SlMYC2 的植株耐寒性最差,证明两者功能互补,缺一不可。
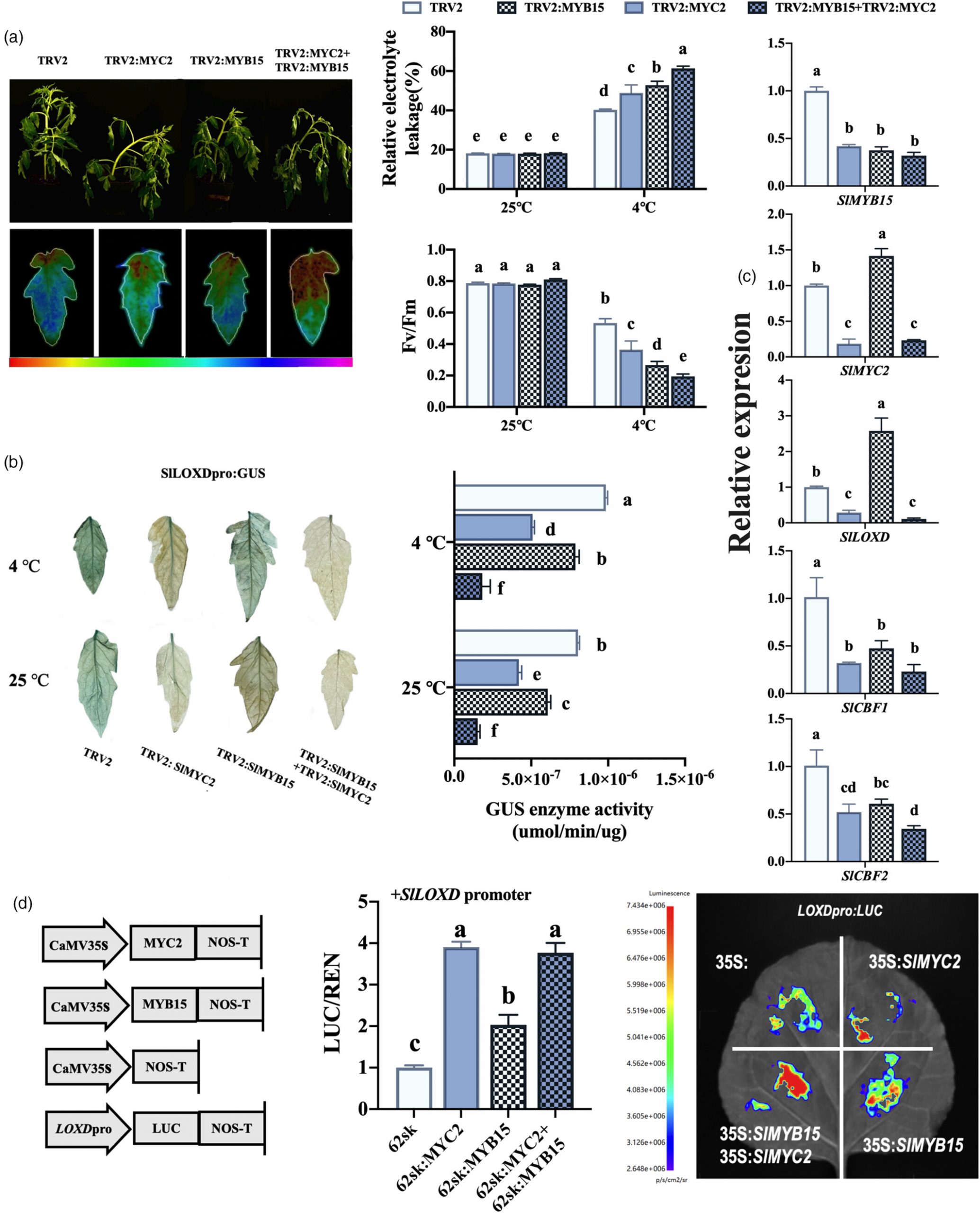
图7 SlMYC2 继续激活 SlLOXD 表达,以提高 SlMYB15 沉默番茄品系的低温耐受性
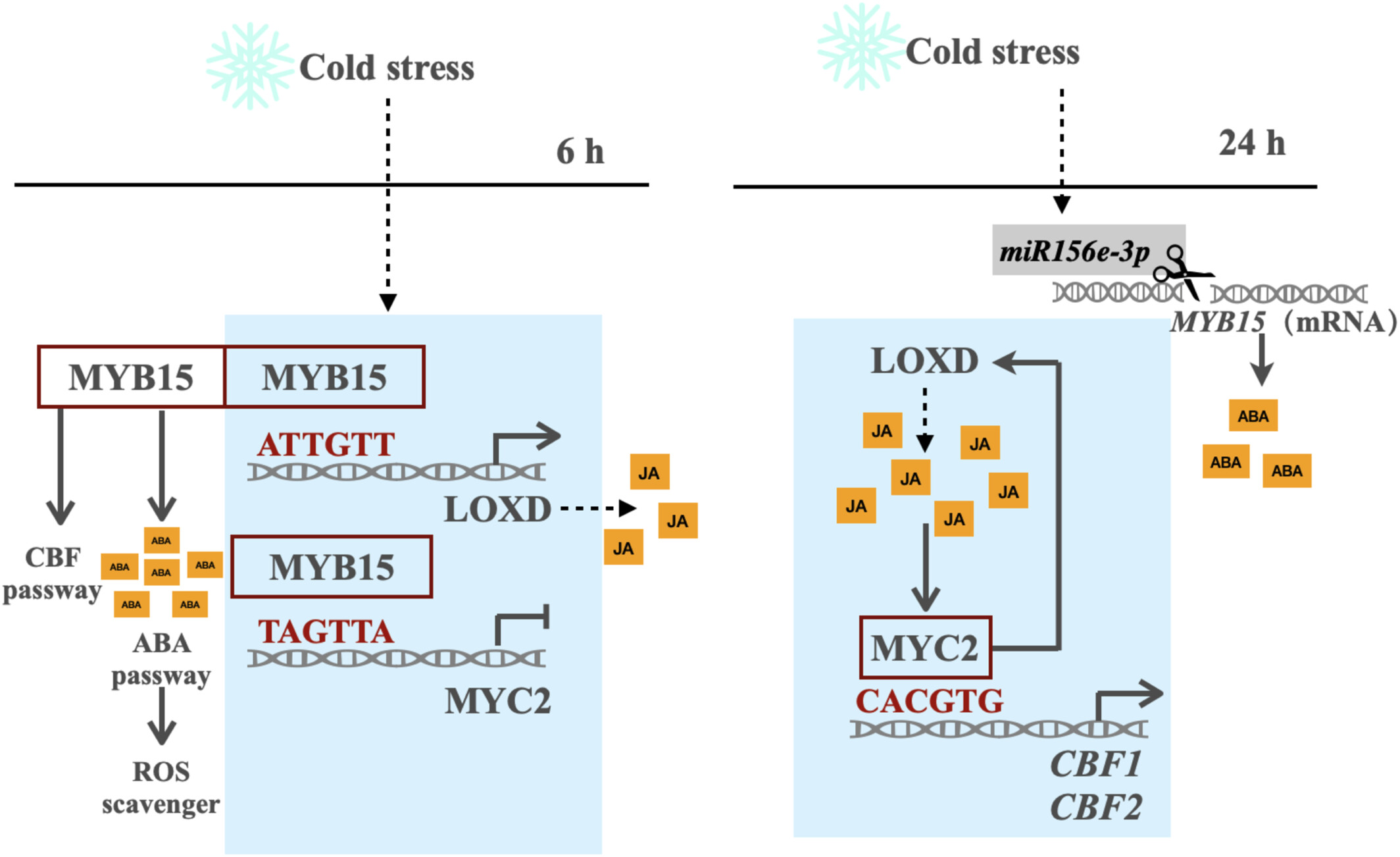
图8 一个功能模型,展示了 SlMYB15 、 SlMYC2 和 SlLOXD 对番茄寒冷胁迫的响应
总结
本文通过多层次实验,清晰解析了番茄冷胁迫中 JA 信号的动态调控网络,证实 SlMYB15 和 SlMYC2 通过时间特异性的协同与接力,借助 SlLOXD 和 SlCBF1/2 实现对耐寒性的精准调控。这一发现不仅深化了植物冷响应机制的理解,也为农业生产中的抗逆改良提供了重要理论依据。


