【JIPB】解锁小麦穗发芽抗性关键:GSK3磷酸化稳定DOG1L4蛋白

【2026春季活动】诺伴春研,提速省心
2026 年 3 月 23 日
【邀您参会】聚焦黄淮海,深耕大豆种,吉农基因邀您共聚涡阳
2026 年 3 月 27 日

本研究发表于《Journal of Integrative Plant Biology》,由四川农业大学团队完成,聚焦小麦生产中亟待解决的穗发芽问题,揭示了GSK3与DOG1L4互作调控小麦种子休眠的全新分子机制,为小麦穗发芽抗性育种提供了关键候选基因和分子靶点,也为植物遗传转化改良作物抗性提供了重要理论支撑。
穗发芽(Pre-harvest sprouting, PHS)是全球小麦生产的重大挑战,收获期持续降雨会削弱种子休眠性,导致种子提前萌发,大幅降低小麦产量和面粉品质。种子休眠性主要由脱落酸(ABA)和赤霉素(GA)的拮抗作用调控,Delay of Germination 1(DOG1)家族是调控种子休眠的核心基因,小麦中TaDOG1Ls也被证实参与种子休眠与萌发调控。此外,油菜素甾醇(BR)信号通路可通过与ABA/GA代谢和信号通路互作,间接调控种子休眠,但BR信号通路核心元件在小麦穗发芽调控中的具体机制尚不明确。研究团队此前已克隆小麦GSK3激酶编码基因TaGSK3,发现其保守基序突变体与小麦矮秆性状相关,本研究进一步探究该基因在小麦穗发芽抗性中的功能及分子机制。
实验思路与方法
本研究以遗传转化技术为核心手段,构建了多个关键转基因材料,系统解析了GSK3调控小麦穗发芽抗性的分子机制:
1、转基因材料构建:利用农杆菌介导的遗传转化技术,在现代小麦品种KN199背景中获得了GSK3功能获得性突变体(GSK3E285K)过表达转基因株系,以及DOG1L4过表达转基因株系,为后续表型与机制研究提供了核心材料。
2、表型鉴定:对上述转基因株系进行穗发芽抗性、种子萌发率、激素处理下的萌发响应等表型分析,明确GSK3与DOG1L4在休眠调控中的作用。
3、蛋白互作与修饰验证:通过Y2H、Pull-down、LCI、BiFC、Co-IP等方法筛选并验证GSK3与DOG1家族成员的互作;利用LC-MS鉴定磷酸化位点,结合点突变体与无细胞蛋白降解实验分析DOG1L4的稳定性。
4、下游通路分析:通过RNA-seq、RT-qPCR、激素测定等分析ABA信号通路相关基因表达变化,结合转基因材料验证GSK3与ABI5的互作与磷酸化调控。
主要研究结果
本研究揭示了小麦中GSK3激酶通过直接与DOG1L4蛋白互作并磷酸化其Ser-294位点,显著增强DOG1L4的蛋白稳定性,从而正向调控种子休眠、提升穗发芽抗性。GSK3功能获得性突变体(GSK3E285K)过表达株系在表型上表现出更强的穗发芽抗性和延迟萌发特性,且这种效应不依赖于GA和ABA的激素含量变化,而是通过稳定DOG1L4实现。进一步研究发现,GSK3还能与ABA信号通路关键转录因子ABI5互作并磷酸化ABI5,协同增强ABA信号响应。激素分析显示,GSK3过表达株系中ABA含量升高、GA和BR含量降低,进一步支持其在多激素调控网络中作为枢纽因子的作用。
该研究还排除了DELLA蛋白在GSK3介导的休眠调控中的作用,明确了DOG1L4是GSK3在休眠调控中的关键下游靶点。
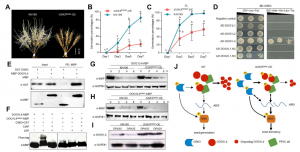
图1. GSK3调控小麦穗发芽抗性
研究意义
- 机制创新:首次将BR信号负调控因子GSK3与种子休眠核心因子DOG1L4联系起来,揭示了GSK3通过磷酸化调控DOG1L4蛋白稳定性的分子机制,拓展了BR信号在种子休眠中的功能认知。
- 育种价值:GSK3功能获得性突变体及其下游靶点DOG1L4为小麦穗发芽抗性育种提供了新的基因资源和分子标记,尤其适合在雨水较多的麦区应用。
- 整合调控网络:研究发现GSK3同时调控DOG1L4和ABI5,形成协同增强休眠的双重机制,为理解激素互作(BR、ABA、GA)在种子成熟与萌发中的整合提供了新视角。
- 技术应用前景:本研究涉及的过表达株系构建、蛋白互作验证、磷酸化位点鉴定等技术体系,为后续通过遗传转化手段改良作物休眠性状提供了可操作的技术路径。
关于我们



