【PBJ】重磅发现!中华根瘤菌鞭毛蛋白“主动激活免疫”调控豆科结瘤

一份来自吉农基因的新春温馨提示与假期安排
2026 年 2 月 9 日
2026实验室扩建计划中!
2026 年 2 月 27 日

豆科植物与根瘤菌的共生固氮,是农业生产中减少氮肥依赖的“天然密码”。长期以来,科学界普遍认为根瘤菌要成功结瘤,必须逃避免植物免疫识别。但华中农业大学曹扬荣团队发表在《Plant Biotechnology Journal》的最新研究,打破了这一传统认知——根瘤菌竟主动激活植物免疫,以此精准调控结瘤过程!这一发现为大豆、苜蓿等豆科作物的高产育种开辟了全新路径。
研究思路与方法
本研究以中华根瘤菌-豆科植物互作体系为研究对象,结合生物信息学分析、分子生物学实验、微生物基因编辑、植物基因编辑(CRISPR/Cas9)、细胞生物学检测、转录组分析、结构生物学模拟等技术,层层解析中华根瘤菌鞭毛蛋白对豆科植物免疫和结瘤的调控机制:
- 生物信息学分析:对中华根瘤菌属鞭毛蛋白序列进行系统分析,鉴定出具有免疫激活潜力的flg22变体。
- 免疫反应实验:通过ROS爆发、MAPK磷酸化、免疫基因表达等实验,验证flg22变体的免疫激活能力。
- 遗传突变体构建:构建中华根瘤菌鞭毛素基因(fliC)突变体及回补株;利用CRISPR-Cas9技术创制大豆GmFLS2a/GmFLS2b单、双突变体及蒺藜苜蓿MtFLS2突变体。
- 表型与分子分析:比较突变体与野生型在结瘤数量、固氮活性、免疫基因表达等方面的差异。
- 结构模拟:利用AlphaFold3模拟flg22与FLS2受体的结合模式。
主要研究结果
1. 关键“信号分子”:flg22Sin-II肽段
研究团队分析了5811个根瘤菌目基因组,鉴定出19160个鞭毛蛋白,发现中华根瘤菌中存在一种特殊肽段——flg22Sin-II。它是鞭毛蛋白的衍生物,仅这一类群具备免疫激活功能,且在中华根瘤菌菌株中广泛分布,是根瘤菌与植物“沟通”的核心信号;而flg22Sin-I和flg22Sin-III则无此功能。
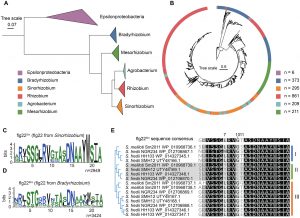
图1. 中华根瘤菌与慢生根瘤菌鞭毛蛋白flg22区域的系统发育与保守性分析
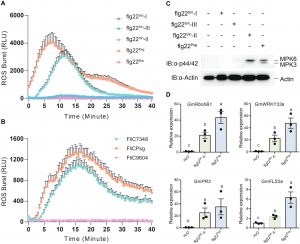
图2. flg22Sin-II 触发大豆的活性氧(ROS)产生
2. flg22免疫识别并负调控结瘤
构建中华根瘤菌HH103(大豆共生菌)、苜蓿中华根瘤菌2011(蒺藜苜蓿共生菌)、中华根瘤菌NGR234(百脉根共生菌)的fliC敲除突变体,接种对应宿主后发现:
- 单敲除fliC不影响根瘤菌的运动能力,排除了运动性对结瘤的干扰;
- 突变体根瘤菌接种后,大豆、蒺藜苜蓿、百脉根的结瘤数显著高于野生型根瘤菌,回补fliC基因后结瘤数恢复至野生型水平;
- 证明含flg22Sin-II的鞭毛蛋白对豆科植物结瘤存在负调控作用。
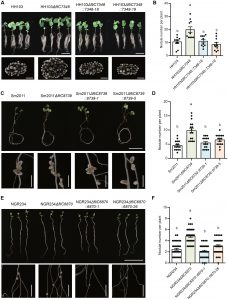
图3. 鞭毛蛋白缺失突变体的结瘤表型
3. 激活免疫的“关键开关”:Tyr-7残基
flg22Sin-II能被豆科植物精准识别,核心依赖其7位的酪氨酸(Tyr-7)。
实验证明:一旦将Tyr-7突变为亮氨酸,该肽段就彻底丧失激活免疫的能力。通过AlphaFold3结构模拟进一步发现,Tyr-7能直接与植物受体GmFLS2a结合,是启动免疫信号的“钥匙”。
4. 宿主介导:FLS2是免疫调控结瘤的“核心媒介”
研究通过CRISPR/Cas9构建大豆和蒺藜苜蓿的FLS2突变体,明确了“flg22Sin-II→FLS2→免疫→结瘤”的调控链。
- 受体功能验证:大豆拥有两个FLS2同源基因(GmFLS2a和 GmFLS2b)。研究发现,它们都能在拟南芥中互补fls2突变体对flg22的响应。更重要的是,利用CRISPR-Cas9技术构建的大豆*GmFLS2a/GmFLS2b*双突变体,对flg22Sin-II和病原菌flg22触发的ROS反应均大幅减弱。
- 共生表型关联:该双突变体在接种野生型中华根瘤菌后,结瘤数显著多于野生型大豆。同样,在蒺藜苜蓿中敲除其MtFLS2基因,也导致了结瘤数增加和固氮酶活性提高。这些结果确凿地证明,植物通过FLS2受体感知根瘤菌的flg22Sin-II,进而激活免疫并抑制共生进程。
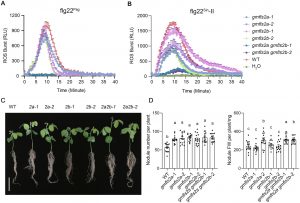
图4. 大豆 GmFLS2a 和 GmFLS2b 参与 flg22 诱导的免疫反应并调控结瘤
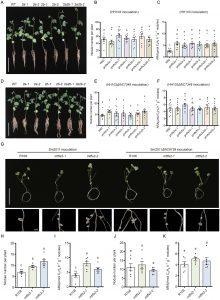
图5. 大豆和蒺藜苜蓿 FLS2 突变体对根瘤菌的结瘤表型
5. 分子调控模型
研究表明:中华根瘤菌侵染时,其鞭毛蛋白中的flg22Sin-II肽段被豆科植物FLS2受体识别;FLS2激活下游免疫信号(ROS 爆发、MPK3/6磷酸化、免疫基因上调);免疫信号抑制共生相关基因的表达,从而限制结瘤数、延迟结瘤时间。 这种“主动激活免疫”的策略,使根瘤菌与植物维持“互利不互损” 的共生稳态,是长期共进化的结果。
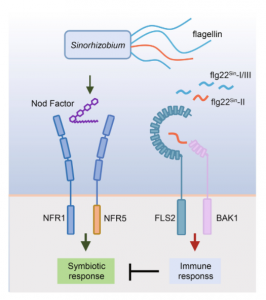
图6. (C) 根瘤菌鞭毛蛋白介导的免疫抑制豆科植物结瘤的调控模型

